Amino acid requirements in developing marine fish
Biologically important molecules have an ancient history
Among the most ancient of molecules, amino acids have existed on Earth longer than life itself. Recent revelations have shown that they are also found in space buried within meteorites. Twenty kinds of amino acids are biologically important. They are all relatively small molecules built from an acid carboxyl group and an alkaline amino group, both attached to a common carbon atom with a side group of variable size and complexity. These variable side groups define the amino acids and give each one its special properties.
Amino acid pools in fish eggs
Amino acids can occur freely dissolved in body fluids (free amino acids, FAAs) or are bound together in long polymerized strings as proteins. Proteins make up about two-thirds of the dry body mass of all animals – including fish. Their synthesis is nutritionally demanding.
A challenge for aquaculturists is providing the large supply of amino acids as well as the energy substrate for this costly synthesis. When growth rate is high, as is the case for fish larvae, the demands are also high.
In the sea, fish embryos and larvae are supplied with a natural diet rich in amino acids. Fish farmers must select artificial diets that mimic this high amino acid availability in order to support the high growth rate of the seedlings.

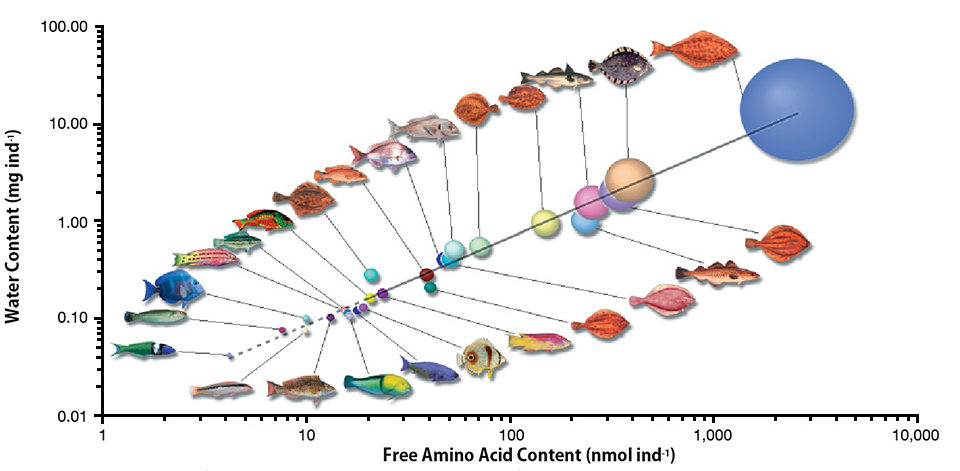
The dry mass of marine fish eggs typically consists of more than 50 percent amino acids in the form of proteins and an additional 15-20 percent as FAAs. Localized to the yolk compartment, the FAA pool has a fairly constant proportion of each amino acid regardless of fish species with pelagic eggs.
The FAA pool size is highly correlated to the water content and size of the eggs (Fig. 1). In fact, the size of the pool is regulated in order to hydrate the egg to greater than 90 percent of its wet mass. These findings have relevance for the functionality of the marine fish egg when spawned into the saline seawater as well as for the fuel preference of the developing larva.
Amino acids as fuel for larvae
The large FAA pool of the newly spawned egg is an important nutritional asset for the growing embryo. It is quickly depleted during a phase of development when major organ differentiation occurs without much growth in body mass.
This occurs in fish eggs without oil globules (type I eggs), where catabolism of FAAs supports about 75 percent of the aerobic energy metabolism during the yolk sac stage, as well as in eggs with oil globules (type II eggs), where the FAAs provide about 50 percent of the necessary energy. During specific developmental windows for species such as Atlantic halibut and Atlantic cod, embryos may even depend on FAAs for 100 percent of the fuel for growth processes.
Further, when FAA pools are depleted, larvae start to derive amino acids from the yolk proteins for growth and fuel requirements, although lipids are also burned with the amino acids (Fig. 2).
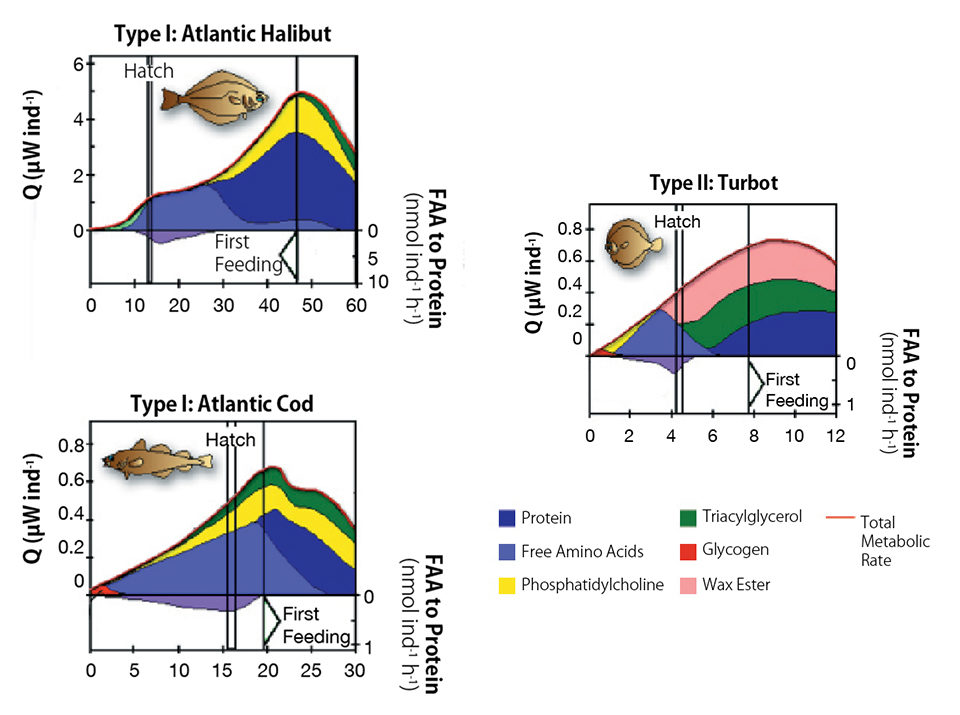
It is noteworthy that in their natural environments, marine fish larvae begin to catch and ingest small planktonic creatures when their yolk reserves are mostly consumed. These creatures are mainly the larvae of other marine animals, such as copepods, worms, mollusks or ciliated microorganisms.
A remarkable feature of these planktonic organisms is that they all contain high concentrations of FAAs in their body tissues as an adaptation to saline marine conditions. Hence, with each ingested plankter, fish larvae receive a new supply of FAA.
During early stages of exogenous feeding, the larval intestine is not yet fully functional with regard to protein degradation, so the fish larvae benefit more from the ingested FAAs than the prey proteins. This notion was supported by experimental findings that showed amino acids to be more efficiently absorbed in larval fish intestines when given a diet of mixed FAAs than when given a diet of proteins.
Osmotic water loss compensation
As hyposmotic animals, marine fish are unique in maintaining their blood concentrations much lower than that of the ambient medium. This implies that water is continuously lost by osmosis from the body fluids, and in order to evade dehydration, the fish must compensate by taking up moisture from the seawater.
This is no small task, but the fish manage by drinking seawater, extracting water and salt ions from the intestinal fluid and excreting the salt ions from the gills and kidneys. The net result is a gain of pure water to replace what is continuously lost so blood concentrations remain stable over time.
Marine fish eggs lack the organs and osmoregulatory mechanisms of grown fish. Still, the fish eggs must compensate for osmotic water loss. Mother fish therefore load eggs with an excess of yolk water before spawning. The yolk FAA pool is part of the mechanism involved in this water transfer to the eggs before spawning.
Origin of egg FAA pool
The large FAA pools of pelagic fish eggs represent a major driving force for water uptake into the yolk compartments. This uptake takes place during oocyte maturation before ovulation and spawning. Special water channels called aquaporins are incorporated into the egg plasma membranes to ease the water flow. The oocytes actually swell five to eight times in volume, and following ovulation and spawning, the eggs float.
The origin of the yolk FAA pool is found in yolk proteins derived from vitellogenin, a special protein produced in the liver and delivered to the oocytes after being transported to the gonads in the blood. There are several kinds of vitellogenin, but yolk proteins derived from the VtgAa type are depolymerized during oocyte maturation, resulting in the build-up of the large FAA pool and the ensuing water uptake and oocyte swelling.
The degradation process is catalyzed by acid hydrolases called cathepsins. The fact that the VtgAa molecules are specifically split explains why the FAA pools produced have such a similar profile in different fish. Other types of yolk proteins derived from VtgAb and VtgC are stored in the yolk as a nutritional amino acid reserve for yolk-sac larvae later in development. How the protein-degradative machinery of the oocytes distinguishes between the different types of vitellogenin is still a mystery.
Evolution of marine fish
The splitting of the VtgAa-derived yolk proteins is a novel but crucial adaptive step that allowed teleost fish to produce eggs that survive in saline seawater. This seems to have allowed the fish to complete their life cycles in the oceanic environment and thus establish themselves as true marine fish.
Based on the fossil record as well as recent analyses of vitellogenin genes and yolk protein sequences, this mechanism emerged about 55 million years ago. The mechanism of oocyte hydration is correlated to the explosive radiation of new fish species that occurred in the oceans at that time. The vast majority of modern marine fish today have pelagic eggs with high water content and large FAA pools.
An important implication of the high egg water content is that the specific gravity of the eggs is lower than that of seawater. This gives the eggs buoyancy and allows them to be spread with the ocean currents, greatly increasing their distribution potential in nature.
For fish farmers, however, the floatation requirements of the newly spawned eggs are an important quality criterion of the produced egg batch. The underlying cellular mechanisms that control the involved acid degradation of yolk proteins appear to have a truly ancient origin, and more basic studies will be needed in order to understand this important aspect of egg quality.
(Editor’s Note: This article was originally published in the March/April 2009 print edition of the Global Aquaculture Advocate.)
Now that you've reached the end of the article ...
… please consider supporting GSA’s mission to advance responsible seafood practices through education, advocacy and third-party assurances. The Advocate aims to document the evolution of responsible seafood practices and share the expansive knowledge of our vast network of contributors.
By becoming a Global Seafood Alliance member, you’re ensuring that all of the pre-competitive work we do through member benefits, resources and events can continue. Individual membership costs just $50 a year.
Not a GSA member? Join us.
Authors
-

Department of Biology
University of Bergen
Bergen High Technology Centre
Postboks 7803
N-5020 Bergen, Norway -
Department of Biology
University of Bergen
Bergen High Technology Centre
Postboks 7803
N-5020 Bergen, Norway
Related Posts

Health & Welfare
Amino acid supplementation reduces protein levels in pangasius diets
Trials show that supplementation with amino acids could reduce protein levels from a typical 28 percent to 23 percent in pangasius diets.

Aquafeeds
A look at corn distillers dried grains with solubles
Corn distillers dried grains with solubles are an economical source of energy, protein and digestible phosphorus to reduce feed costs and fishmeal usage.

Aquafeeds
A look at protease enzymes in crustacean nutrition
Food digestion involves digestive enzymes to break down polymeric macromolecules and facilitate nutrient absorption. Enzyme supplementation in aquafeeds is a major alternative to improve feed quality and nutrient digestibility, gut health, compensate digestive enzymes when needed, and may also improve immune responses.

Aquafeeds
A new nutrient for aquaculture, from microbes that consume carbon waste
Biotechnology firm NovoNutrients aims to produce a line of nutraceutical aquafeed additives as well as a bulk feed ingredient that can supplement fishmeal. Its process includes feeding carbon dioxide from industrial gas to a “microbial consortium” starring hydrogen-oxidizing bacteria.