Scope for further optimization of selective breeding in aquaculture species
Capitalize on additive genetic effects

Genetic improvement programs are key for the development of better performing stocks in aquaculture. Their crucial importance for better utilization of limited feed and land resources is widely recognized, and implementation of well-designed programs is strongly advocated as reflected in FAO’s Code of Conduct of Responsible Fisheries. This article briefly describes the general structure of family-based selection programs, and identifies areas where recent developments in genetics and related fields may further increase their efficiency for aquaculture species.
Improvement through effective selection
The application of quantitative genetics and selection theory in aquaculture dates back several decades, and numerous selective breeding programs have been implemented. Many of these programs have been based on individual selection for improved growth rate. Although this selection method can be highly effective for this trait, individual selection for growth rate has often led to poor results, most likely due to inadequate hatchery management procedures resulting in severe inbreeding problems.
Today, the number of more complex, family-based selection programs is increasing fast, and at present more then 25 are in operation worldwide. Their main advantage compared to individual selection programs is their flexibility and superior efficiency for improving traits of intermediate to low heritability and traits that cannot be recorded on live animals at a controlled and predefined rate of inbreeding in the target populations.
Aquaculture species ideal for improvement work
The high fecundity and short generation interval of many aquaculture species make them excellent candidates for large-scale selection programs (Table 1). Very high reproduction capacity facilitates strong selection intensity, and many traits of primary importance in aquaculture productions show high levels of additive genetic variation. Moreover, separate collection of eggs and milt and external fertilization in most candidate species opens a wide range of alternative family group designs.
Rye, Generation interval and fecundity, Table 1
| Species | Generation time | No. eggs/kg body weight |
|---|---|---|
| Atlantic salmon | 4 | 1,600 |
| Rainbow trout | 3 | 1,800 |
| Atlantic halibut | 4-6 | 40,000 |
| Atlantic cod | 3 | 200,000 |
| Sea bass | 3 | 300,000 |
| Sea bream | 2-4 | 500,000 |
| Turbot | 2-3 | > 1 million |
| Pacific white shrimp | 1 | 1-1.5 million* |
| Black tiger shrimp | 1 | 1-2 million* |
| Giant freshwater prawn | 1 | – |
Provided appropriate tagging methods, family-based breeding designs can be applied. Combined family and within-family selection increases the selection efficiency for traits of low heritability (e.g., survival), and enables selection for traits that can only be recorded through destructive tests (e.g., carcass quality), or disturbing tests that cannot be undertaken in the breeding nucleus (e.g., disease challenge). For further refinement, selection may be combined with designs for crossing stocks or genetic lines, thus utilizing non-additive as well as additive genetic components of genetic variation.
Cost-efficient programs
High fecundity allows concentration of resources in one or a few breeding centers, keeping operational costs low. The genetic gain obtained in the nucleus is typically disseminated throughout the target industry with a minimum time lag through a level or collaborating multiplier units, often through commercial hatcheries. High fecundity also makes it possible to select for a broad and long-term breeding objective in the breeding nucleus, while at the same time selecting for one or few traits for the production of grow-out animals. In this way, the genetic level for the traits of principal importance to the producers will be higher in the growout animals, when compared with the animals selected for reproduction of the breeding nucleus. This possibility reduces the conflict between long- and short-term objectives often experienced in livestock.
The “Engine”
The breeding nucleus operations constitute the “engine” of a selective breeding program, and provide accumulated selection responses for the target traits if efficient selection is applied and if the rate of inbreeding accumulation in the nucleus population is controlled. For family-based programs, the nucleus operations (Fig. 1) typically involve production and incubation of a number of full- and half-sib families according to the chosen mating design; separate rearing of families till the animals reach a minimum size which allows physical tagging; tagging followed by pooling of animals from different families; and distribution of samples from all families for performance testing and as breeding candidates. Performance testing for production traits (e.g. growth, survival, age at sexual maturation, and carcass quality traits) are usually carried out in commercial production systems, while disturbing tests required for recording traits like disease resistance are mostly carried out in facilities dedicated to that specific purpose.
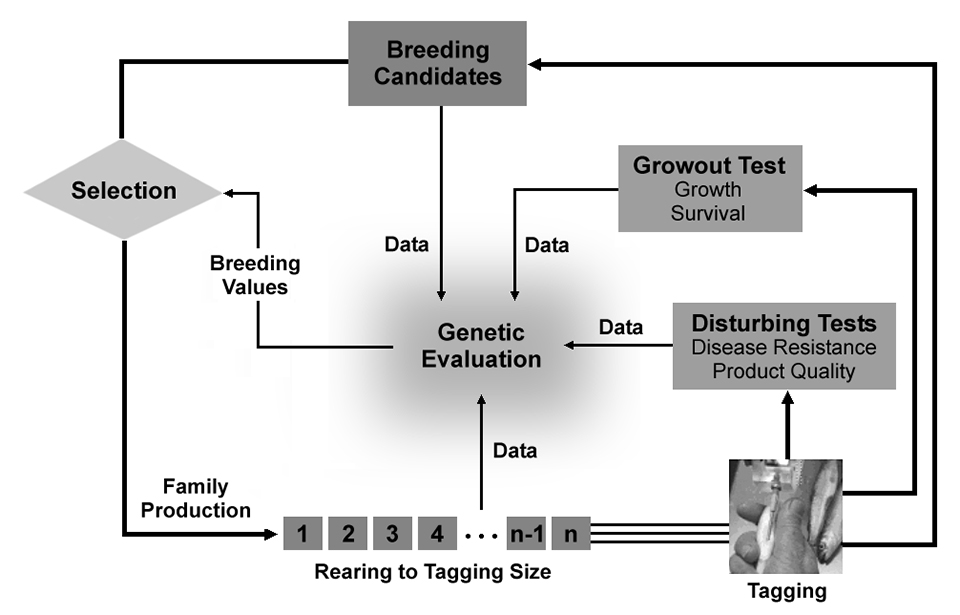
Data recorded at various stages in the life cycle of the animals serve as base for the genetic evaluations, and the estimated breeding values are used for the final selection of breeders to be used for reproducing the breeding nucleus, and selection of breeders for production of eggs or fry at the multiplier units. Given that the effect of genotype by environment interaction is low (i.e., resulting in fairly consistent ranking of families across production environments), use of performance data from different commercial farm environments secure selection of breeders that are robust to differences in environmental conditions.
The distinctive reproductive characteristics of aquaculture species allow for accurate estimation of breeding values and for high selection intensities, and therefore high short-term selection responses. However, high selection intensities and accuracies may result in a rapid accumulation of inbreeding that can jeopardize further genetic improvement, reduce fitness and performance, and produce highly variable responses. Implementation of measures for restricting inbreeding is therefore essential in programs for aquaculture species.
Scope for further optimization
Development work in various relevant fields may further enhance the efficiency of applied genetic improvement programs.
Customized genetic material
While controlling the accumulation of inbreeding in the breeding nucleus is vital for sustained long-term selection response, inbreeding is not an issue when producing grow-out animals since these are not to be reproduced. Therefore, the cost of maintaining broodstock for dissemination purposes is usually low. Furthermore, effective dissemination may facilitate production of highly customized genetic material for different production environments or market segments with specific product requirements. In most selection programs currently in operation, this potential is not exploited.
Improved recording technology
Presently, several traits of economic importance cannot be recorded on live breeding candidates due to the lack of adequate recording technology. If this problem could be overcome, it would facilitate increased selection intensities for these traits and consequently allow more rapid genetic gains. Alternatively, the same rate of improvement could be obtained with the use of fewer resources in terms of the number of families and individuals that needs to be tested.
Improved models for genetic evaluations
Non-additive genetic effects are not accounted for in current procedures used for routine breeding value estimations, but recent advances in the development of effective algorithms for large data-sets now makes this feasible. Extended models including non-additive genetic effects are expected to give improved accuracy for additive genetic merits, and thus increased selection responses. Non-additive genetic effects may also be exploited, particularly at the multiplier level, through specific mate allocation.
In particular, optimizing the genetic evaluation models used for disease resistance traits should be focused. Selection for improved resistance is mostly based on breeding values estimated by fitting a linear model to full-sib family challenge test data, even if the phenotypic expression of disease resistance is non-linear. Implementation of non-linear evaluation models for disease resistance traits allow more efficient use of the available data, and may increase the selection efficiency and thus the response. Use of appropriate non-linear evaluation models may also lower the number of animals needed in the challenge test and shorten the required testing period.
Extended use of DNA fingerprinting?
Sophisticated genetic techniques may become increasingly more valuable as supplementary tools in selective breeding. Tracking genetic relationships among animals in the breeding nucleus is of vital importance in any family based-program. Use of hyper-variable DNA elements (microsatellites) allows families and even individuals within families to be identified by DNA fingerprinting without the need for physically tagging the individuals. This may allow for pooling of different families at early stages, and thus reduce the need for multiunit facilities and the potential problem of environmental effects common to members of the same family.
On the other hand, large-scale use of DNA fingerprinting for this purpose carries substantial and recurrent costs, since all breeding candidates as well as test animals must be DNA-typed. At present, the cost issue is effectively blocking widespread application of DNA fingerprinting for identification purposes in large-scale improvement programs in aquaculture. In addition, tracking of genetic relationships by DNA markers carries risk related to representation of individual families in the breeding population. Unbalanced population structures lead to decreased selection intensities and reduced selection responses.
Marker-assisted selection?
In theory, superior breeding programs can be implemented by inclusion of information on mapped production genes in the evaluation of breeding candidates. The development of dense genetic maps makes it possible to localize regions, or loci, associated with variation in quantitative traits (QTLs), and may facilitate marker-assisted selection. However, the optimal use of molecular information needs to be determined, in particular when selection is simultaneously made for several traits. In addition to this, most – if not all – traits of economic importance in aquaculture are multi-factorial and thus influenced by a large number of genes which individually have only marginal impact on the phenotypic expression of the trait. The additional gain to be expected from inclusion of marker information in improvement programs is therefore limited.
Conclusion
Well-designed breeding programs capitalizing on additive genetic effects have lead to remarkable improvements in several fish and shellfish species. Still, further optimization of the programs is needed in order to adapt the full range of quantitative and molecular genetics tools to these new species.
(Editor’s Note: This article was originally published in the February 2003 print edition of the Global Aquaculture Advocate.)
Now that you've reached the end of the article ...
… please consider supporting GSA’s mission to advance responsible seafood practices through education, advocacy and third-party assurances. The Advocate aims to document the evolution of responsible seafood practices and share the expansive knowledge of our vast network of contributors.
By becoming a Global Seafood Alliance member, you’re ensuring that all of the pre-competitive work we do through member benefits, resources and events can continue. Individual membership costs just $50 a year.
Not a GSA member? Join us.
Authors
-

Akvaforsk Genetics Center
Ås N-6600 Sunndalsøra, Norway -
Institute of Aquaculture Research (AKVAFORSK)
N-1430 Ås Norway
Related Posts

Health & Welfare
Black tiger domestication, selective breeding advance in Australia
Using clear-water tank systems, CSIRO and a collaborating farm have advanced the domestication of black tiger stocks in Australia.

Health & Welfare
10 paths to low productivity and profitability with tilapia in sub-Saharan Africa
Tilapia culture in sub-Saharan Africa suffers from low productivity and profitability. A comprehensive management approach is needed to address the root causes.

Intelligence
Adding value to tilapia to tap into U.S. market
New markets for tilapia and expansion of existing ones can be created by planning and implementing properly designed geographic strategies to meet discriminating consumer preferences. Low labor costs in most producing countries promotes value-adding by the production of fresh fillets.

Aquafeeds
Alternative lipids spare fish oil in rainbow trout feeds
Alternative lipids have achieved varied success in ensuring adequate growth and fatty acid composition in fillets. The authors evaluated rainbow trout raised on diets containing fish oil or a blend of fish and standard or modified lipids varying in fatty acid composition.