Microminerals are important feed components
Organic trace elements more effective than inorganic forms
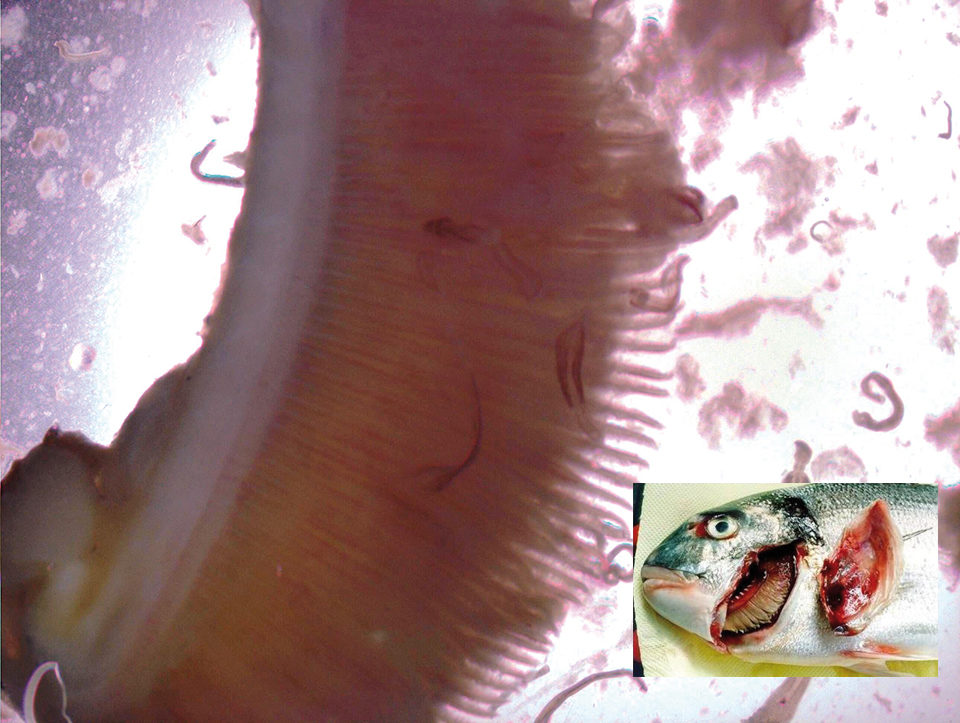
The roles of microminerals or trace elements in basic metabolic functions, with the exception of osmoregulation, are the same for aquatic and terrestrial animals. These microminerals, such as chromium, cobalt, copper, iodine, iron, manganese, molybdenum, selenium and zinc, are required in small quantities and participate in a wide variety of biochemical processes (Table 1).
Nengas, Microminerals, Table 1
| Micromineral | Functions |
|---|---|
| Chromium | Chromium is associated with the glucose tolerance factor, an organometallic molecule that potentiates the action of insulin, important in carbohydrate metabolism. |
| Cobalt | Cobalt and intestinal bacteria are important in the synthesis of vitamin B12. |
| Copper | Copper participates in hematopoiesis, in copper dependent metalloenzymes responsible for oxidation reduction and in the absorption and metabolism of iron. |
| Iodine | Iodine is an essential component of thyroid hormones important in regulating the metabolic rate of all body processes. It has roles in thermoregulation, intermediary metabolism, reproduction, growth and development, hematopoiesis, and circulation and neuromuscular functioning. |
| Iron | Iron is essential for the production of haemoglobin, myoglobin, cytochromes and many other enzyme systems. Iron is one of the primary metals involved in lipid oxidation. |
| Manganese | Manganese is a cofactor or component in several key enzymes responsible for bone formation, in urea synthesis, amino acid metabolism and glucose oxidation. |
| Molybdenum | Molybdenum is a cofactor in xanthine oxidase. |
| Selenium | Selenium is an essential component of the enzyme family glutathione peroxidase, which protects cells from deleterious effects of peroxides. Selenium acts along with vitamin E to function as a biological antioxidant to protect polyunsaturated phospholipids in cellular and subcellular membranes from peroxidative damage. |
| Zinc | Zinc functions as a cofactor in several enzyme systems and is a component of a number of metalloenzymes required for normal growth, development and function. |
They are involved in cellular metabolism, formation of skeletal structures, maintenance of colloidal systems, regulation of acid-base equilibrium and other physiological functions. They are important components of hormones and enzymes, and serve as cofactors and/or activators of a variety of enzymes. However, in comparison to the terrestrial animal industry, our understanding of the micromineral requirements of fish is often limited.
Diets
Global aquaculture in developed countries is increasingly represented by intensive farming practices that use nutritionally complete feeds as the sole source of nutrition. Traditionally, fishmeal has been a major component of these diets. Other feedstuffs of animal origin, such as meat and bone meal, poultry by-product meal and feather meal, are also relatively rich in minerals. However, due to consumers’ preferences and legislation related to recent health scares in the terrestrial animal sector, they are excluded from aquafeeds in some regions.
Diet formulations have seen radical structural changes in the last five years due to economic and sustainability considerations that have demanded a reduction in fishmeal and fish oil usage and their replacement by more sustainable, alternative plant protein and vegetable oil sources.
Modified and unmodified plant protein sources, such as soymeal, legume seeds, oil seed cakes, leaf meals, leaf protein concentrates and root tuber meals, are now widely used, as well as single-celled proteins such as yeast and algae produced using fermentation techniques.
Dietary concerns
Major concerns in using alternative protein sources in aquafeeds include their nutrient profiles, poor mineral sources and the presence of anti-nutritional factors that can affect micromineral bioavailability. Increasing dietary plant proteins results in an increase in indigestible components, fiber and non-starch polysaccharides, and an associated increase in levels of phytic acid and its negative interactions in mineral availability. Further, the available phosphorous, phospholipids and general digestibility of the protein content are also affected. Dietary zinc availability and absorption are reduced in the presence of phytates.
How do fish obtain minerals?
The determination of mineral dietary requirements for aquatic animals is challenging due to their ability, unlike terrestrial animals, to absorb minerals directly from their external aquatic environment in addition to those supplied through their diets.
Freshwater fish absorb salts through their gills. Combined with a low body surface, this permeability enables the fish to maintain blood ion levels at higher concentrations than in the external environment. Marine fish loose water through all permeable surfaces and therefore have to replace the lost water osmotically by drinking seawater, allowing absorption of ions and water through the gut into the blood.
Fish are able to regulate their body concentrations of microminerals at near-normal levels by excreting excessive micromineral intakes in urine and feces. Traditionally, inorganic forms of microminerals are used as supplements in feeds, but their poor bioavailability and discharge in uneaten food and feces directly influence the aquatic environment, particularly the sediments under cages.
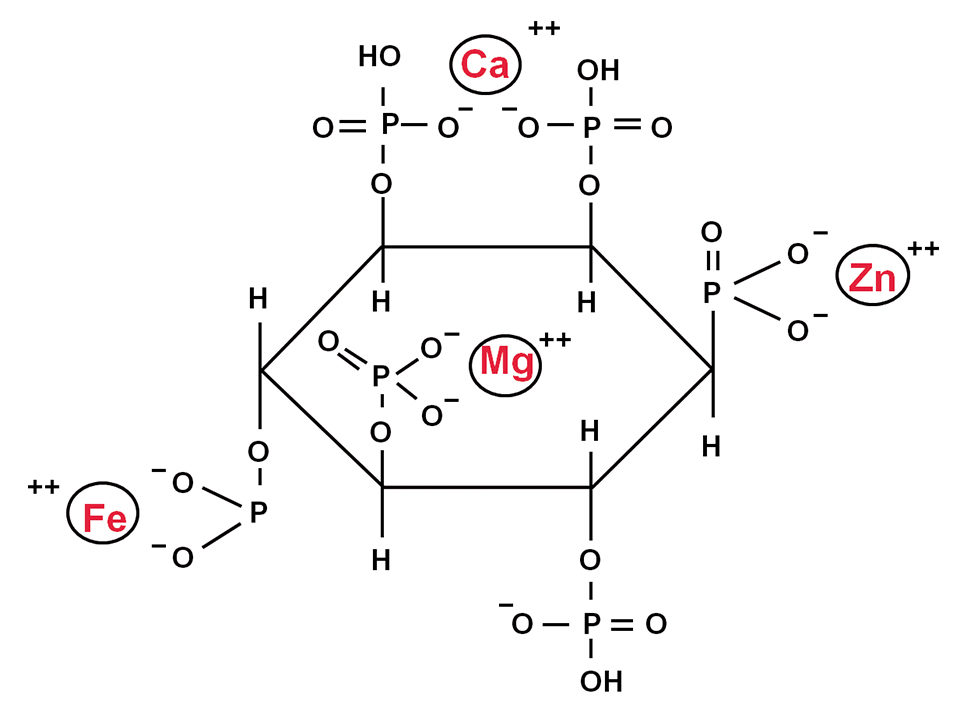
Mineral bioavailability, storage
The bioavailability and tissue storage of trace minerals are of primary importance in their nutritional performance. Minerals chelated to organic molecules have higher bioavailability than their corresponding inorganic forms and interact less with each other in the digestive tract. Amino acid chelates of cobalt, manganese and zinc are more readily available than their inorganic salts, while organic chelate zinc methionine has been estimated as three times more potent than the inorganic sulphate.
The organic forms can greatly enhance the absorption of an element by either releasing the element or being absorbed as an intact chelate. Chelated minerals are less sensitive to the inhibitory action of other compounds because of their reduced solubility in water. The increasing bioavailability and effectiveness of organic micromineral forms can dramatically reduce the required inputs and decrease waste discharges to the environment.
When the dietary supply of microminerals exceeds requirements, many tissues act as storage reserves, with organic micromineral forms more effectively accumulated than inorganic forms. Fish retain microminerals in soft tissues such as muscle, liver and bone, and measurements of these are used as indicators of the nutritional levels of practical diets.
Stress, disease resistance
In intensive culture, fluctuating environmental and water quality conditions combined with practical husbandry practices can result in the development of stress situations that affect the physiology and micromineral requirements for the species cultured. Indeed, as reported in Aquaculture in 2009, Sebastien Rider demonstrated that chronic physical stress in rainbow trout increased selenium utilization, as indicated by significantly decreased tissue reserves of selenium and increased glutathione peroxidase activity. He also noted that during stressful situations, the selenium status was more effectively maintained by an organic form of selenium than by the inorganic form.
Stress conditions can also affect the osmotic and ionic regulation in gills. In freshwater fish, this can result in passive ion losses and water uptake. In seawater species, it can increase passive ion influxes and water loss, while inhibiting active ion exchanges in both.
Organic chelated iron has been used to alleviate anemia symptoms and mortality caused by microcotyle parasitic infections and their interactions with haematocrit blood cells in sea bream.
Immune defense
Interest in microminerals has especially been strengthened by the importance of their roles in immune defense and antioxidative protection. The significance of selenium in fish nutrition is markedly growing along with the intensification of the industry.
Similar to other feed industries, organic selenium has been found more effective in fish nutrition than its inorganic forms. In the immune system, selenium is particularly important, as it is incorporated into active selenoenzymes as selenocysteine. These protect the phagocytic cells of the innate immune system – one of the best characterized aspects of the essential nature of selenium in disease defense.
Nutritionally complete diets are necessary in culture situations and, therefore, micronutrients must be supplied in adequate levels in the prepared diets to support optimal growth and production efficiency. This is particularly true in intensive situations, where immunocompetence and disease resistance can be substantially compromised by deficiencies of various nutrients, especially vitamins and microminerals.
Dietary supplementation of some of these micronutrients in excess of minimum requirement levels has been shown to have a positive influence on immunity, as well as resistance to and recovery from disease.
While the inorganic micromineral forms are adequate to prevent complete deficiency in most species, there is a clear trend for all diet ingredients for humans and animals, especially production animals, toward more biologically available forms.
(Editor’s Note: This article was originally published in the May/June 2012 print edition of the Global Aquaculture Advocate.)
Author
-

Hellenic Centre for Marine Research
Institute of Aquaculture
Agios Kosmas
Elleniko 16777 Greece
Related Posts

Aquafeeds
A look at phospholipids in aquafeeds
Phospholipids are the major constituents of cell membranes and are vital to the normal function of every cell and organ. The inclusion of phospholipids in aquafeeds ensures increased growth, better survival and stress resistance, and prevention of skeletal deformities of larval and juvenile stages of fish and shellfish species.

Innovation & Investment
A review of unit processes in RAS systems
Since un-ionized ammonia-nitrogen and nitrite-nitrogen are toxic to most finfish, controlling their concentrations in culture tanks is a primary objective in the design of recirculating aquaculture systems.
Intelligence
An engineer’s design for a classroom aquaculture-aquaponics system
An aquaponics teaching system was designed, built and operated by students at the University of Arizona, integrating its operation and management into the educational curriculum. This engineering design will require minimum maintenance and will last years.

Aquafeeds
Aquaculture Exchange: Andrew Jackson, IFFO
Aquaculture remains dependent on fishmeal and fish oil, crucial marine ingredients in aquafeeds, particularly at key life stages. Andrew Jackson, technical director at IFFO and one of the world’s foremost fishmeal experts, tells the Advocate that the two industries can coexist well into the future if properly managed.