Four AHPND strains identified on Latin American shrimp farms
EMS update: First report on pathogenic Vibrio campbellii strains

Acute hepatopancreatic necrosis disease (AHPND), also commonly known as Early Mortality Syndrome (EMS), is a serious, major global disease of farmed shrimp that causes hepatopancreas dysfunction and secondary Vibrio infections and can result in 100 percent mortality in the animals’ juvenile stages.
The disease started in China in 2010 and has currently spread to aquaculture facilities in Vietnam, Malaysia, Thailand, the Philippines, Mexico and much of Latin America. Losses from EMS have been estimated to exceed $1 billion per year.
The literature reports AHPND as caused by a unique strain of Vibrio parahaemolyticus, an ubiquitous bacterium that colonizes uneaten feed and organic sludge in culture pond bottoms, and also the chitinous shrimp molts and the animals’ stomach linings, and is transmitted horizontally from shrimp to shrimp and vertically from spawner to egg. Here we present the first report describing the pathogenicity of non-V. parahaemolyticus AHPND strains.
Bacterial isolation and AHPND identification
Four Vibrio strains (16-902/1, 16-903/1, 16-904/1 and 16-905/1) were isolated from either stomachs of diseased shrimp or sediment samples from AHPND-affected farms in Latin America during 2016. Bacterial identifications were carried out using 16S rRNA sequencing and Vibrio-specific PCR assays targeting hly gene. These bacteria were grown on TSB+ (Tryptic soy broth plus 2 percent NaCl) at 28-29 degrees-C with gentle shaking (100 rpm) and further tested for AHPND PCR assays with primers targeting pirABvp.

By the PCR assays, these 4 strains were identified to V. campbellii by 16S rRNA sequence analysis and hly gene PCR. These V. campbellii strains had both pirAvp and pirBvp genes (Table).
AHPND pathogenicity bioassays
We determined pathogenicity of a representative AHPND V. campbellii strain (16-904/1) by shrimp bioassays. Three 90-L tanks were filled with artificial seawater (salinity of 25 ppt and temperature at 28 degrees-C), and specific-pathogen free (SPF) P. vannamei (N=20, weights: 2.0 g) were stocked in each tank. For the infection, each strain was cultured in TSB+ overnight with gentle shaking (100 rpm) to reach 1×109 CFU/mL, and shrimp were exposed the bacterial culture by immersion at a concentration of 2×105 CFU/mL water. Shrimp were monitored for mortality every 12 hr. Also, AHPND V. parahaemolyticus strains, 13-028/A3 (Vietnam strain, 2013) was tested as a positive control.
During the assays, no mortality was observed during the five-day bioassay period in a negative control. However, the pirABvp-containing V. campbellii strain 16-904/1 caused AHPND in P. vannamei and all the shrimp became moribund or died within two days, same as a positive control (Table).
| Strain | Origin (year) | VpPirAB (a) | Pathogenicity (b) |
|---|---|---|---|
| AHPND V. campbellii 16-902/1 | Latin America (2016) | Pos | Not determined |
| AHPND V. campbellii 16-903/1 | Latin America (2016) | Pos | Not determined |
| AHPND V. campbellii 16-904/1 | Latin America (2016) | Pos | Pathogenic |
| AHPND V. campbellii 16-905/1 | Latin America (2016) | Pos | Not determined |
| AHPND V. parahaemolyticus 13-028/A3 | Vietnam (2013) | Pos | Pathogenic |
Histopathology
For histopathology, shrimp were fixed in Davidson’s alcohol-formalin-acetic acid (AFA), and analyzed for histopathology. The Davidson’s AFA-fixed samples were processed, embedded in paraffin, and sectioned (4 μm thick) in accordance with standard methods. After staining with hematoxylin and eosin (H&E), the sections were analyzed by light microscopy.
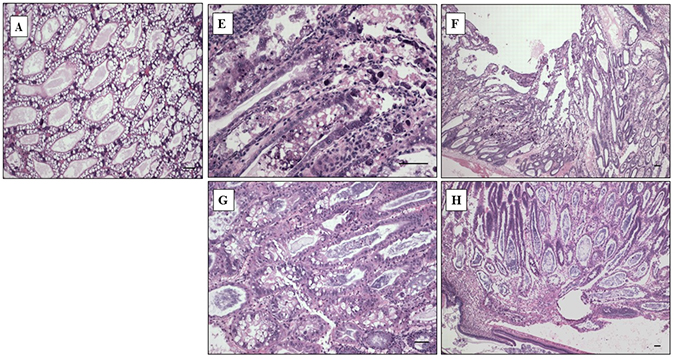
Negative control (non-AHPND V. parahaemolyticus) showed the normal appearance of the hepatopancreas (Figure A). However, shrimp challenged by pirABvp-containing V. campbellii strain 16-904/1 were confirmed to AHPND by histological examination (Figure E&F). The histological signs showed a severe necrosis of hepatopancreas tubules with a massive sloughing of epithelial cells, and hemocytic infiltrations in severely affected tubules in the shrimp. The mortality and the histological signs were even higher than that of positive controls (AHPND V. parahaemolyticus) (Figure G&H).
Perspectives
Our findings of 4 pirABvp-containing V. campbellii strains confirm that these strains are pathogenic to shrimp in bioassays and cause AHPND. This is the first report describing the pathogenicity of non-V. parahaemolyticus AHPND strains. These findings confirmed that pirABvp are the virulence genes responsible for AHPND and also indicate a horizontal gene transfer between different bacterial species (from V. parahaemolyticus to V. campbellii) in natural ponds.
Now that you've reached the end of the article ...
… please consider supporting GSA’s mission to advance responsible seafood practices through education, advocacy and third-party assurances. The Advocate aims to document the evolution of responsible seafood practices and share the expansive knowledge of our vast network of contributors.
By becoming a Global Seafood Alliance member, you’re ensuring that all of the pre-competitive work we do through member benefits, resources and events can continue. Individual membership costs just $50 a year.
Not a GSA member? Join us.
Authors
-

Corresponding author
CJ CheilJedang Feed & Livestock Research Institute,
CJ Blossom Park,
Suwon-si, Gyeonggi-do, Korea
-

School of Animal and Comparative Biomedical Sciences
University of Arizona
Tucson, AZ 85721 USA
-

School of Animal and Comparative Biomedical Sciences
University of Arizona
Tucson, AZ 85721 USA -
School of Animal and Comparative Biomedical Sciences
University of Arizona,
Tucson, Arizona, USA
-
Department of Public Administration,
Yonsei University
Seoul, Korea
Related Posts

Health & Welfare
Genetic variation for resistance to WSS, AHPND in Pacific white shrimp
Selection for disease resistance has been used in breeding farm animals and can be a viable option to deal with white spot syndrome and acute hepatopancreatic necrosis disease in commercial shrimp culture. In trials, heritability for AHPND resistance was low, while that for WSS was moderate.

Health & Welfare
Limited decomposition enhances PCR detection of AHPND Vibrio in shrimp
A study confirmed the utility for improved polymerase chain reaction (PCR) detection of the Vibrio bacteria that cause acute hepatopancreatic necrosis disease (AHPND) in asymptomatic shrimp by permitting the shrimp to expire and decompose for several hours prior to preservation and PCR processing.

Health & Welfare
PCR methods characterize AHPND V.p isolates
In analyzing the plasmid sequence from the whole genome sequences of AHPND V. parahaemolyticus (V.p) isolates, researchers identified a clear geographical variation within the plasmid, and developed PCR methods to characterize AHPND V.p isolates as either Mexico-type or Southeast Asia-type.

Health & Welfare
A holistic management approach to EMS
Early Mortality Syndrome has devastated farmed shrimp in Asia and Latin America. With better understanding of the pathogen and the development and improvement of novel strategies, shrimp farmers are now able to better manage the disease.

Health & Welfare
Probiotics benefit Pacific white shrimp challenged with AHPND
A study was conducted to measure the effects of commercial probiotics on Pacific white shrimp in a standardized AHPND challenge model under controlled laboratory conditions. Results show that the probiotics treatments by themselves have beneficial effects, such as higher survival and histological signs of hepatopancreas regeneration.
Health & Welfare
Probióticos benefician al camarón blanco del Pacífico desafiado con AHPND
Un estudio fue llevado a cabo para medir los efectos de probióticos comerciales en camarón blanco del Pacífico en un modelo de desafío a AHPND estandarizado bajo condiciones controladas de laboratorio. Los resultados muestran que los tratamientos de probióticos por sí mismos tienen efectos beneficiosos, como una mayor supervivencia y signos histológicos de regeneración de hepatopáncreas.